At chromosome level
Sometimes whole sections of text in our instruction manual are repeated or omitted, even sections that consist of millions of letters. We call these CNVs or Copy Number Variations. They can be inherited or can occur 'de novo'.
Continue reading below the image.
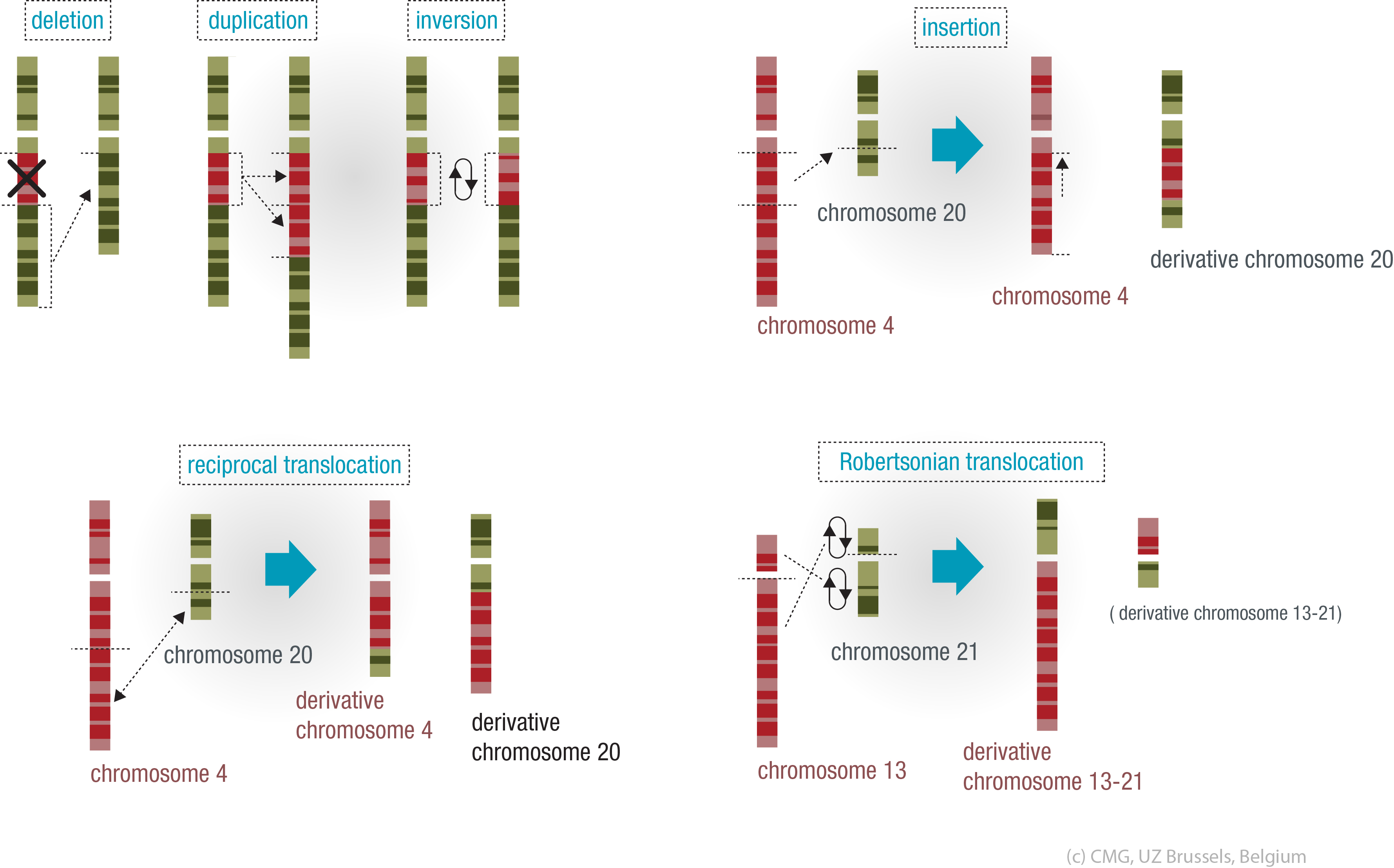
Deletion
If genetic material is lost, we call this a deletion. Depending on the amount we call these macrodeletions or microdeletions, but the consequences can be severe in both cases.
In the case of a microdeletion, genes or parts of genes may be missing. One example of a disorder that is the result of a microdeletion (on chromosome 22) is Velo-cardio-facial syndrome.
Sometimes a deletion is so large that a whole chromosome is missing. This is called a 'monosomy'. In almost all these cases the fetus is not viable and will die spontaneously. Monosomy of the X chromosome is the only one that allows the fertilised egg cell to develop into an embryo and continue to develop as a human being. In that case the baby will be a girl with only 45 chromosomes (Turner syndrome).
Duplication
If an error during cell division results in the doubling of a chromosome or part of a chromosome, we call this duplication. This can occur in both autosomes and sex chromosomes.
If a whole chromosome is doubled, we call this a trisomy. People with a trisomy - quite logically - have 47 rather than 46 chromosomes. This usually results in severe abnormalities. As a result the embryo is not viable and a spontaneous miscarriage occurs. The three viable autosomal trisomies are trisomy 21 (Down's syndrome), trisomy 18 (Edwards syndrome) and trisomy 13 (Patau syndrome).
Sex chromosomes that are duplicated may also result in viable variants. Thus, there are girls with three X chromosomes (Triple X syndrome) and boys with two Y chromosomes and an X chromosome or two X chromosomes and a Y chromosome (Klinefelter's syndrome).
Sometimes only part of a chromosome is duplicated: we call this a partial duplication. The duplicated part may be joined to the chromosome where it belongs, but it may also be joined to another one (duplication with translocation).
Inversion
In an inversion, part of the genetic material is exchanged within a single chromosome. As long as no genetic material is lost as a result, this will usually not result in an inherited disorder affecting the carrier of the inversion. Her egg cells or his sperm cells may, however, acquire an imbalance between the chromosomes during cell division.
Translocation
In a translocation, part of one chromosome is joined to another (non-homologous) chromosome.
We call this a balanced translocation if no genetic material is lost or duplicated. The simple shifting of genetic material does not usually cause a problem for the person who is a carrier, but he or she will have an increased risk of having descendants with an unbalanced translocation.
In an unbalanced translocation the descendant will inherit the affected chromosome with either more or less genetic material than the parent's balanced translocation. Since the genetic code is changed as a result, this usually causes problems for the carrier.
Two types of translocations are known:
- Reciprocal translocations, in which part of the a chromosome is exchanged
- Robertsonian, whole-arm translocations
Marker chromosomes
A marker chromosome is extra chromosomal material that cannot be clearly identified or characterised using conventional banding techniques. It is actually a partial trisomy that originates from one or more of the 24 different chromosomes.
Marker chromosomes can be inherited (approximately 30%) or may arise de novo (approximately 70%).
The impact of a marker chromosome completely depends on its contents. If the marker chromosome mostly contains inactive genetic material, its effect will be minimal.
Often marker chromosomes are not inherited because they are small chromosomes and are therefore lost during cell division.
One of the marker chromosomes that occurs quite frequently is a reversed duplication of chromosome 15.
Fragile sites and break points
At certain specific, fragile positions on chromosomes in metaphase, 'gaps' may occur which can lead to spontaneous break points during cell division.
There are more than 120 fragile positions that are currently known in the human genome. These can be classified as 'general' or 'rare' on the basis of the frequency with which they occur.
General fragile positions occur in (almost all) normal individuals. They often consist of regions where DNA is repeated (repeats). The most extensively studied form of rare fragile position is FRAXA, which results in the Fragile X syndrome, a form of inherited mental retardation caused by expansion of a CGG repeat.